BY LETTER
Xenobiology and Natural History of Tohul
Science > Biology > Evolutionary Biology
Sophonts > Xenosophonts > To'ul'hs
Science > Biology > Xenobiology
under constructionSophonts > Xenosophonts > To'ul'hs
Science > Biology > Xenobiology
 Image from Luke Campbell | |
| Beneath the thick clouds of Tohul Prime the surface is dim, but lightning briefly bathes the landscape in bright light. | |
Table of Contents
A Brief Overview of Modern Tohulian Biota
- Surface and Lower Atmosphere
- Middle and Upper Atmosphere
- List of Prominent Tohul Indigenous Lifeforms
Geological and Evolutionary History of Tohul
- Prototohulian Eon: 3500-2280 mya
- Chaotohulian Period: 3500-2880 mya
- Pelatohulian Period: 2880-2280 mya
- Eotohulian Eon: 2280-1200 mya
- Cryotohulian Eon: 1200-1000 mya
- Campotohulian Eon: 1000-400 mya
- Tartarean Eon: 400-300 mya
- Caelosilvanic Eon: 300 mya-present
Tohul is a hot terrestrial world with a dense, wet atmosphere, with a surface atmospheric pressure nearly 20 times that of Earth. The planet supports a rich biosphere, more complex and diverse than that of any other Garden World of its subtype discovered to date. Multicellular life-forms can be found in the photic zone of the upper atmosphere, where conditions are fairly similar to those of Earth at sea level, as well as in the lower atmosphere and surface environments, where little light reaches and ambient temperature and pressure conditions reach extremes that are intolerable to most naturally-evolved Terragen organisms.
Overview of Tohulian Biology
As with Earth and many other Garden Worlds, life on Tohul appears to have arisen just a single time, with all modern Tohulian organisms tracing their ancestry to a local abiogenesis event which took place around 1.8 billion years BT. As is also the case with most Garden Worlds, the majority of Tohulian taxa are unicellular prokaryote-equivalents; these comprise several domain-level groups broadly similar to Terragen bacteria and archaea, as well as another which blends characteristics of bacteria and viruses. Multicellular eukaryote-equivalents have evolved at least three times, each from a different set of prokaryote-equivalent ancestors. Numerous classes of viruses and other mobile genetic elements are also present, parasitizing and propagating within life-forms at all levels of complexity.Life on Tohul is for the most part strikingly similar to naturally-evolved Terragen life on a biochemical level, consisting of aqueous cells encapsulated in phospholipid bilayers, which utilize DNA and RNA for genetic information storage and transfer, and proteins for a wide variety of structural and biochemical functions. Even the chirality of the proteins and nucleic acids found in Tohulian life happens to be the same as those found in Earth's native life, with proteins being levochiral (left-handed) and nucleic acids being dextrochiral (right-handed). However, the sets of amino acids and nucleobases found in Tohulian proteins and nucleic acids only partially overlap with those found in Earth life.
The surface temperature on Tohul ranges from 361-408 degrees Kelvin (88-135 degrees Celsius), far outside the tolerance of virtually all native Earth organisms aside from certain extremophilic microbes. All life on Tohul's surface exhibits a set of cellular-level adaptations which enables them to maintain homeostasis at these extreme conditions; many of these mirror traits seen in hyperthermophiles on Earth, such as rigidified cell membranes with a high proportion of unsaturated fatty acids, ultra-stable proteins which utilize extensive ionic bonding and disulfide cross-linking to prevent unfolding under heat stress, and elaborate mechanisms for repairing and refolding denatured biomolecules. All Tohulian organisms are also adapted to the relatively high proportion of sulfur, tin, mercury, and antimony compounds that are present in Tohul's atmosphere and crust due to the planet's high metallicity. Unlike many Garden Worlds, in which life can extend for kilometers below the surface, Tohul's subsurface crust is too hot to support any form of life at depth, as the surface temperatures are already close to the limits beyond which protein-based life cannot function.
The three largest "superkingdoms" of eukaryote-equivalents on Tohul, respectively labeled Phlegethonia, Asphodelia, and Elysia by Terragen xenobiologists, are each adapted to a particular temperature regime, although their ranges overlap to some degree. Each superkingdom contains both unicellular protist-like life-forms as well as multicellular organisms broadly comparable to animals, plants, or fungi.
The hyperthermophilic Phlegethonia are primarily found within the oceanic basins and continental surfaces, and prefer temperatures between 373-408 degrees Kelvin (100-135 degrees Celsius). This superkingdom contains several kingdoms of multicellular life, the most prominent being the fungus-like Thermomycota (commonly known as "fungoids"), the terrestrial sponge-like Pulvispongia (commonly known as "land-sponges"), the non-photosynthetic but otherwise plant-like Putrephyta (commonly known as "putrephytes"), and the animal-like Tohulanimalia.
Of the various kingdoms of Phlegethonia, the Thermomycota comprise the greatest proportion of the surface biomass, growing wherever any significant volume of sediment and organic matter accumulates. Like Terragen fungi, they consist of a mass of mycelium-like filaments that extract nutrients from their substrate, often visible as a moldy fuzz on the soil, and also produce transient fruiting bodies that disperse spore-like gametes. These fruiting bodies commonly resemble Earth mushrooms like puffballs and stinkhorns, and may range in size from a few centimeters to over a meter in diameter. A few lineages produce balloon-like structures filled with methane gas to aid in spore dispersal; these along with various other surface organisms form the foundation of nascent cloudforest mats, which helps to circulate surface nutrients into the upper atmosphere.
Although less abundant than the fungoids, members of the Putrephyta represent the largest sessile life-forms of the surface landscape. Like fungoids, putrephytes are also saprotrophic organisms, but they more closely resemble non-photosynthetic vascular plants than Terragen fungi, having complex root-like structures belowground and permanent freestanding stems or trunks above the surface. Their aboveground portions are usually simple, unbranched poles or stalks bearing clusters of reproductive organs at the top, with the smaller species resembling the stalks and seed-heads of Earth grasses, and the larger tree-equivalents somewhat resembling massive paintbrushes set on end; as no putrephytes are photosynthetic, they lack any structures resembling leaves. The brush of reproductive structures at the top of the pole or stem contains male cone-like organs that produce pollen, and female cone- or drupe-analogs that enclose seeds. Most species are wind-pollinated, but some have evolved the equivalent of flowers, which attract pollinators using distinctive scents, bioluminescent organs, or distinctive structures that rattle, buzz or whistle in the breeze. The larger varieties of putrephyte grow in "forests" wherever there is sufficient water and protection from the wind, and the continental uplands are covered in "grasslands" of lower-growing species.
The Pulvispongia somewhat resemble terrestrial versions of ancient archaeocyathid sponges from Earth's Cambrian Era, consisting of a porous tubular body anchored by a holdfast. The majority of species are shaped like narrow vases, either alone or as a cluster, though some form a thin crust of polyp-like forms instead. Like Earth sponges, they consist mostly of skeletal elements, with only a thin layer of living tissue covering their internal support structure. They may be found growing wherever water and airborne dust are sufficient to support life, usually on substrates that are too poor in organics to support putrephytes; some species may also be found on the trunks of the larger putrephytes, and in a few cases form symbiotic associations with them.
The kingdom Tohulanimalia includes a dozen or more phyla of organisms broadly equivalent to Earth animals. Notable among these are the Caenovermidae, whose members resemble annelids and molluscs and primarily burrow into the sediment, the Chthonibrachidae, a phylum of suspension feeders which resemble echinoderms, corals and bryozoans, and the Tohulichordates, a phylum of relatively large and active motile organisms that feed on saprophytes or other animal-equivalents. This last phylum includes the Tohultetrapoda, a class of animal-equivalents with four limbs, internal skeletons and highly-developed nervous systems, which encompasses the To'ul'hs and their relatives. Unlike Earth tetrapods, members of this phylum lack distinct heads, with the central nervous system being localized within the center of mass at the junction of the limbs; their ancestors were four-armed crinoid-like suspension feeders, and as a result all members of this class exhibit some degree of radial fourfold symmetry. In some lineages, the ventrally-located jaws are extended into a forward-projecting feeding appendage, and the skin folds between the hind legs may be elaborated into a tail. Arthropod-like phyla are also numerous, as on many Garden Worlds, and fill a wide variety of niches ranging from burrowers in the sediment, suspension-feeders, and consumers of sessile saprophytes or smaller animal-equivalents. Few motile organisms weigh more than 500 kilograms, perhaps due to the low oxygen content of the atmosphere, but since the air is so dense many are capable of gliding or powered flight, and in many cases these may reach surprisingly large sizes; floating planktonic forms are also common, with several groups utilizing buoyancy chambers filled with lighter-than-air gas.
In addition to the Phlegethonia, the surface and lower-to-middle atmospheric cloud decks are inhabited by the Asphodelia, which favor temperatures around 358 degrees Kelvin (85 degrees Celsius). On the surface, these are primarily found on the highest mountains of the continental plateaus, where temperatures are relatively cool. Most members of this superkingdom take on relatively simple crustose forms that extract nutrients from soil and decomposing organic matter, or aeroplanktonic organisms that subsist on airborne particulates; there are several lineages in the latter category that have evolved to utilize the dim light in the middle cloud decks for photosynthesis. The largest organisms within the Asphodelia somewhat resemble land-living versions of Earth coelenterates, and live as suspension feeders and passively-hunting predators that either float in the dense lower atmosphere using gas-filled buoyancy chambers or are anchored to the ground in a manner similar to Earth corals or anemones.
The third superkingdom of eukaryote-equivalents, Elysia, is adapted to a temperature range more broadly compatible with native Earth life, centered around 308 degrees Kelvin (35 degrees Celsius), and is found exclusively in the photic zone of the upper atmosphere, though many have spore or seed forms that can survive the temperatures of the lower atmosphere and surface environments. This superkingdom predominantly consists of the photosynthetic organisms that form the trophic foundation of all surface ecosystems on Tohul, along with several relatively small kingdoms of animal- and fungus-equivalents. These groups collectively form the bulk of Tohul's middle- and upper-atmosphere aeroplankton, typically adopting unicellular or microscopic multicellular forms, but they also include macroscopic green-pigmented phototrophic plant-equivalents that make up much of the biomass of the upper-atmosphere "cloudforest" ecosystems. There are also five additional phyla of floating algae-equivalents among the Elysia, which utilize red, brown, yellow, purple, and blue photosynthetic or accessory pigments. These "sky-algae" maintain their buoyancy using bladders of oxygen gas, although some use nitrogen instead in order to avoid combustion from lightning strikes. All Elysia primarily utilize lipids rather than as carbohydrates for energy storage; this is in part an adaptation to reduce weight, as lipids are much more energy dense for their mass.
A Brief Overview of Modern Tohulian Biota
Tohul's biosphere can be roughly divided into two regimes: the hot, dim, high-pressure surface and lower atmosphere, and the cooler, brightly-lit lower-pressure middle and upper atmosphere.Surface and Lower Atmosphere
Tohul's surface is very dim, with around 99.9% of all light from H'tat'sa'thoss being blocked by the atmosphere's thick cloud cover, dense swarms of aerial micro- and macrobiota, and the ubiquitous haze of slowly-descending biological particulate matter in the lower atmospheric levels. While many motile animal-equivalents on the surface have some degree of light-sensing capability, with some even utilizing bioluminescence for communication and display, the ever-present clouds of dust and organic particulates severely limits visibility, and thus the vast majority of organisms on the surface are functionally blind by nearbaseline human standards.The largest and most active animal-equivalents in most surface ecosystems are members of the Tohultetrapoda, the class which encompasses To'ul'hs and other closely related organisms. Notable among these are the Shur'rooss'hur (Lututherium spp.), relatively large and robust detritivores which are commonly used by the To'ul'hs as beasts of burden, and the H'shass'thee ("Spockbats", Deinoparvichiropteryx spp.), small flying predators whose limbs have specialized into four clawed, membranous wings.
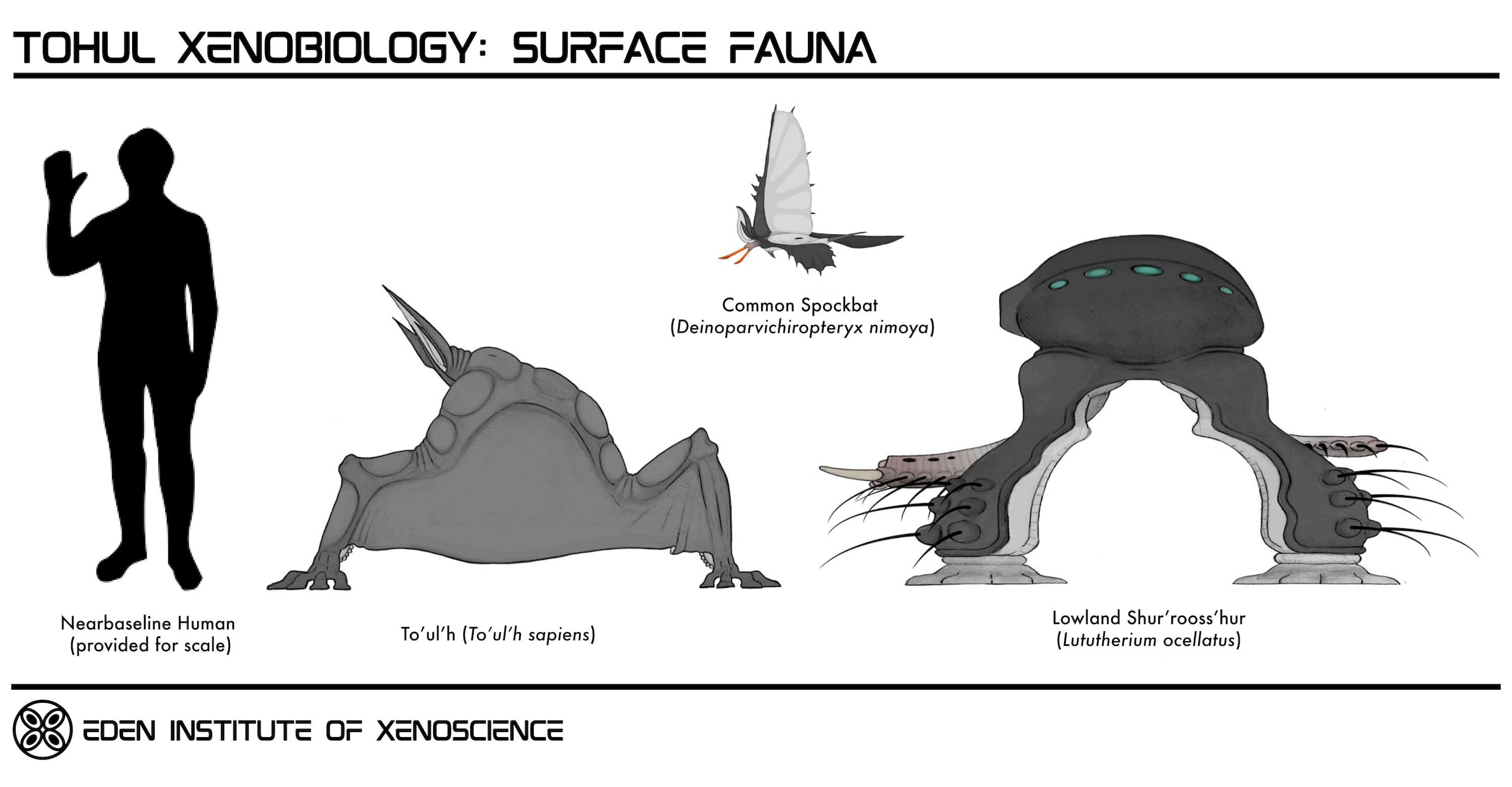 Image from Andrew P. |
 Image from Arik |
 Image from Steve Bowers | |
| Tho'rahl'shothan land sponges, made visible on the dark surface of To'ul'h Prime by their phosphorescent glow | |
The primary source of nutrients in the oceans is a constant rain of "marine snow," originating both from the atmospheric ecosystem and runoff from terrestrial environments. The seabed at nearly all depths is densely populated with a wide variety of sessile detritus-feeders, generally adopting morphologies resembling Terragen kelp and deep-ocean sea anemones, with arrays of tentacles and basket-like filtering structures designed to catch particles of suspended biomass or small swimming organisms. Living among this tangled profusion of benthic organisms are a huge diversity of marine animal-equivalents, commonly known to Terragens as "blindfish," so named because nearly all species completely lack any kind of visual sense. Instead, they primarily rely on electrical and mechanosensory organs, similar to those found in Terragen sharks and other cartilaginous fish, to navigate their surroundings and detect prey or predators.
Two of the more common types of pelagic blindfish are the "Tesla eels" (Fulgurichthys spp.), hagfish-like scavengers capable of generating powerful electrical currents for defense against predators, and the "zapswords" (Electrogladius spp.), slow-moving predators with broad, rippling fins and elongated "bills" that can similarly discharge powerful electric shocks to stun or kill prey. Both these and many other species of blindfish are regularly consumed by To'ul'hs, although insulating protective gear must be worn by To'ul'h "fishermen" to ensure their safety. Owing to Tohul's thick atmosphere, a relatively large number of aquatic clades are also capable of both swimming and aerial gliding or even powered flight, using fins or siphons for locomotion both below and above the water's surface; two of the most common groups in this category are the "flying squids" (Volateuthoidea), which resemble four-armed cephalopods with multiple pairs of wing-like fins, and the "mantoids" (Xenobatoidea), so named for their passing resemblance to Terragen manta rays.
Middle and Upper Atmosphere
 Image from Dangerous Safety |
Among the most numerous animal-equivalents of the photic zone are the relatively simple medusa-like and balloon-like floaters of the Asphodelia; there are also many forms of Phlegethonia which have adapted to the colder temperatures, including arthropod- and tetrapod-equivalents within the Tohulanimalia. Many of these are capable of gliding or powered flight, while others are flightless and resident on or within cloudforests.
The Sagan Drift is a region of mostly clear airspace in the middle troposphere, spanning from 15-20 kilometers above sea level across the entire planet. Many of Tohul's largest and most famous skyfauna may be found in this region.
 Image from Steve Bowers |
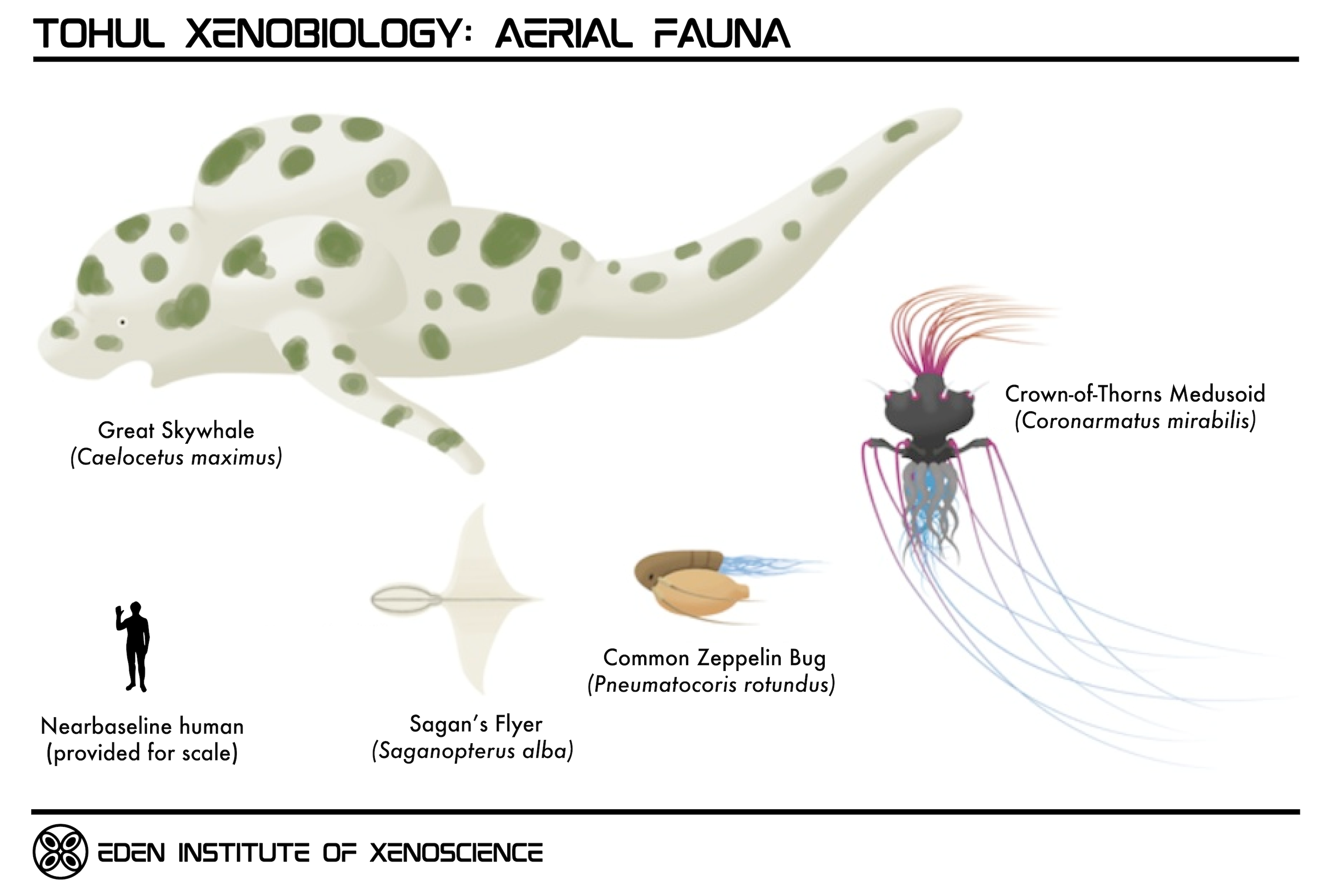 Image from Andrew P. | |
| Some of the larger skyfauna which thrive in the atmosphere of To'ul'h Prime | |
Skywhales are by far the largest organisms on Tohul, with the Great Skywhale reaching up to two hundred meters in length. Like aquatic Terragen whales, skywhales are filter feeders, feeding upon clouds of aeroplankton as they travel in massive herds or "pods" of several thousand individuals. They are preyed upon by "dragons" (Tohuldraco venator), large pack-hunting apex predators distantly related to other skyfish, which utilize their specialized saw-like mouthparts to carve off chunks of skywhale flesh; thanks to the compartmentalized nature of skywhale lift chambers, a pack of dragons can feed on a skywhale for hours before it loses buoyancy and sinks into the depths below.
 Image from RAP 2000 |
A cloudforest begins on the surface, where hyperthermophilic microbes form mat-like communities interwoven with fungoids, putrephytes and other sessile saprophytic organisms. As the microbes break down organic matter, they release methane and hydrogen as metabolic by-products, forming bubbles of trapped gas beneath the filamentous surface of the mat; in many cases, various species of fungoid will also produce gas-filled fruiting bodies as part of their natural life cycle, as they can disperse spores and seeds widely from an elevation. Eventually, the gasses collectively produced by these organisms generate enough lift that the mat breaks free of the surface, carrying with it a large volume of soil.
As the mat rises, various species adapted to cooler temperatures become more abundant, and the hyperthermophilic surface life becomes dormant or produces spores. The increasing light levels result in a gradual increase in photosynthetic life-forms, until somewhere in the middle-to-upper atmosphere photosynthesis becomes the primary source of energy; at this stage, the mat is considered a mature cloudforest. Land-sponges and other cold-tolerant sessile filter-feeders carried up from the surface continue to gather nutrients from atmospheric particulates, and the surviving saprophytes continue to recycle the available nutrients within the mass of the forest. The overall appearance and ecological characteristics of a cloudforest depends on the specific groups of organisms that make it up, along with its location in the atmosphere.
The largest cloudforests may be many hectares in extent, growing either through the expansion of the original mat or by merging with other mats as they drift through the atmosphere. They are inhabited by a bewildering array of macroscopic life-forms, consisting of a mix of Elysia, Asphodelia and various lineages of Phlegethonia that have adjusted to the lower temperatures of the middle-to-upper troposphere. Among the most notable animal-equivalents in the latter category are the "flying monkeys" (Xenolemuridae spp.), eight-limbed creatures with prehensile tails, well-developed eyes and four sets of patagia stretched between their limbs, each of which terminates in a dextrous set of digits. Countless species of arthropod-equivalents can also be found crawling on or flying around the cloudforests, many of which are preyed upon by small, fast-moving skyfish or passively hunting medusoids.
Many species of flying organisms nest in cloudforests and hunt or forage in the zones below, bringing up new micronutrients; some sessile cloudforest organisms actually grow specialized structures that serve as nesting or roosting spots for flying animal-equivalents, so as to benefit from the fertilizer gained from their excretions. The most complex Elysia have a motile animal-like stage of their life cycle with a primitive analog of a nervous system and wing-like appendages that enable powered flight, which seek out and colonize floating mats in the lower depths and draw them upwards into the photic zone.
Over time, the size and mass of the cloudforest increases as phototrophs continue to fix atmospheric carbon dioxide and nitrogen, in most cases eventually reaching a point where it fissions into multiple smaller "islands" of floating life. As micronutrients are gradually lost and the captured soil of the cloudforest becomes less fertile, its constituent life-forms begin to die off or leave in search of better conditions, and the cloudforest gradually decays and disintegrates. The pieces sink or fall back to the surface, carrying with them spores and seeds which grow and sprout again on the surface or in other rising cloudforests. This process plays an important role in the nutrient recycling of Tohul's planetary biosphere, and is a primary contributor to the planet's overall extraordinary fertility and biodiversity.
List of Prominent Tohul Indigenous Lifeforms:
- Blindfish
- Crown-of-Thorns Medusoid
- Tohulian Dragon
- Flying Monkey
- Locust-Beetles
- Sagan's Flyer
- Shur'rooss'shur
- Skyfish
- Skywhale
- Spockbat
- To'ul'h
- Tho'rahl'shothan
- Zeppelin Bug
Geological and Evolutionary History of Tohul
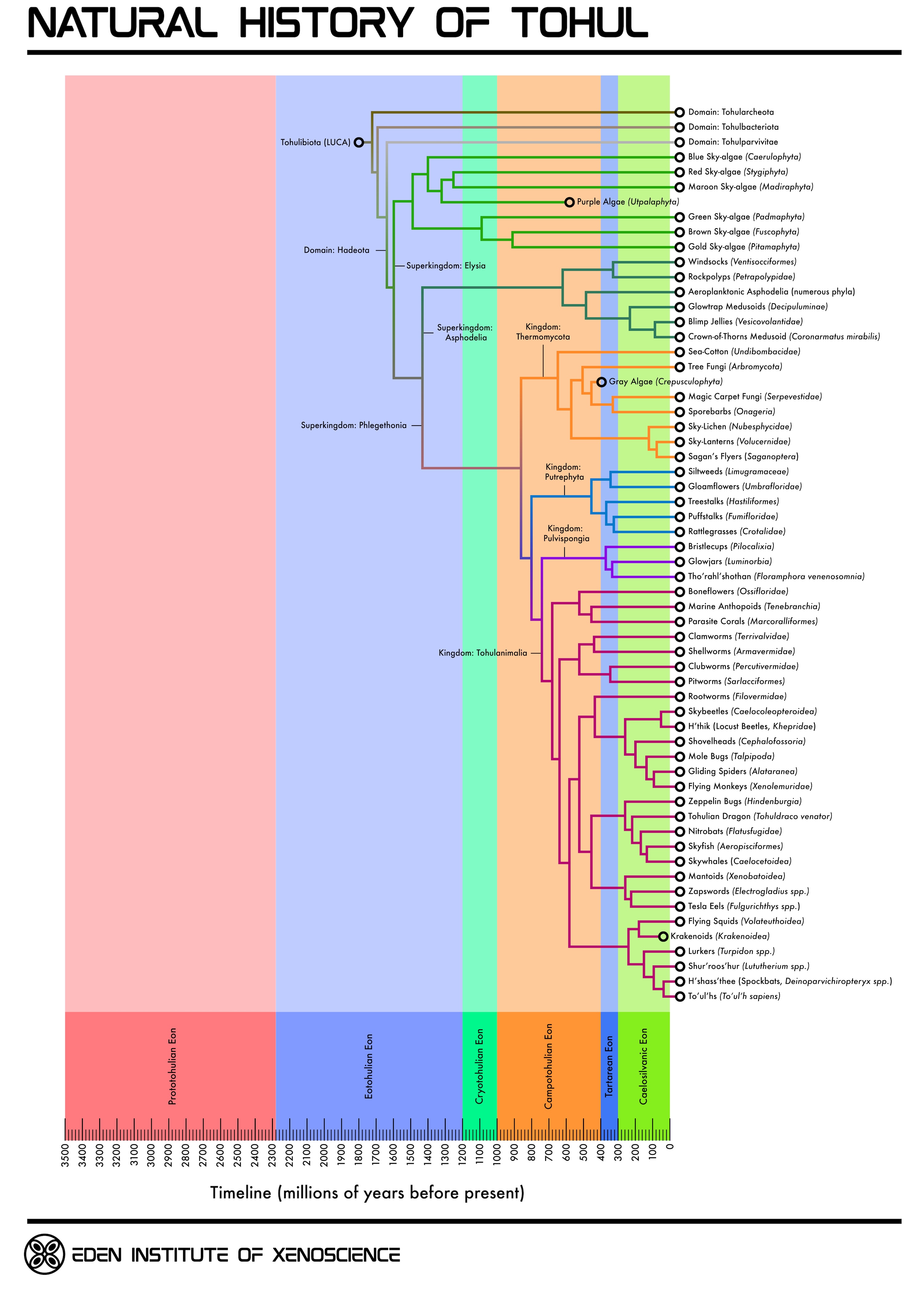 Image from Andrew P. |
Prototohulian Eon: 3500-2280 mya
The Prototohulian Eon encompasses the formation of Tohul and its oceans. It is divided into two periods:Chaotohulian Period: 3500-2880 mya
Tohul forms from a protoplanetary disk surrounding H'tat'sa'thoss. The atmosphere is initially very thick and composed of hydrogen, carbon monoxide, and water vapor; over time, the carbon monoxide is rapidly oxidized by water into carbon dioxide, and atmospheric hydrogen is lost to space.Pelatohulian Period: 2880-2280 mya
A short-lived period during which Tohul had oceans of supercritical carbon dioxide. Surface temperatures during this period never drop below 673 degrees Kelvin (400 degrees Celsius). Surface atmospheric pressure decreases from ~440 to ~215 bar as water vapor condenses and enters the oceans, taking dissolved carbon dioxide with it.Eotohulian Eon: 2280-1200 mya
The Eotohulian Eon on Tohul was broadly similar to the Hadean Era on Earth. The atmosphere at the start of the Eotohulian was initially almost entirely carbon dioxide and was quite thick, around 170 bar dry and 215 bar wet. This dropped to only 60 bar dry / 70 bar wet by the end of the Eotohulian. Nitrogen and methane gradually began to make up more of the atmosphere as carbon dioxide became sequestered into the planetary interior. Surface temperatures during this eon ranged from 453-513 degrees Kelvin (180-240 degrees Celsius).Life first appeared on Tohul during the Eotohulian, and diversified into four domains: Tohulbacteriota, Tohularcheota, Tohulparvivitae, and Hadeota. The Hadeota further diverged into the three superkingdoms which would later give rise to Tohul's modern groups of eukaryote-equivalents: Elysia (predominately photosynthetic organisms adapted to moderate/low temperatures), Asphodelia (mesothermophiles adapted to the temperatures found at higher surface elevations) and Phlegethonia (chemoheterotrophic hyperthermophiles adapted to the extremely hot ocean temperatures). The Elysia subsequently gave rise to the kingdom known as Narakabiota, which diverged into two lineages of photo-autotrophic microbes, the green-pigmented Padmaphyta and the purple-pigmented Utpalaphyta, as well as a group of chemo-heterotrophs, the Avicibiota.
Cryotohulian Eon: 1200-1000 mya
Life on Tohul continued to diversify during this eon. Various groups of photosynthetic microbes, mainly within the Utpalaphyta and their offshoot taxa, the red-pigmented Stygiphyta and blue-pigmented Caerulophyta, began to produce substantial amounts of oxygen as a metabolic by-product of photosynthesis. The oxygen produced by these organisms was toxic both to themselves and other contemporary microbes, much as was the case on Earth when the first cyanobacteria evolved. This resulted in a bottleneck that shaped life on Tohul into its modern metabolic categories, with Phlegethonia and Asphodelia developing aerobic metabolisms, and the Elysia tending toward facultative anaerobic lifestyles.Increased oxygen resulted in the oxidation of atmospheric methane into water and carbon dioxide, reducing the planetary greenhouse effect and causing global temperatures to drop. This resulted in the formation of ice clouds in the upper troposphere, which reflected sunlight and caused temperatures to fall even faster. Life in the upper atmosphere struggled to adapt to this change, but life on the surface (which remained at about 363 degrees Celsius) was relatively unaffected.
Utpalaphyta and Padmaphyta specialized into different niches around this time. Utpalaphyta grew in large blooms on the surface of the oceans, while the Padmaphyta lived at deeper levels, reproducing more slowly but using a more efficient form of photosynthesis. Meanwhile, the Phlegethonia grew to dominate the lower water column, and members of the Avicibiota derived energy from chemicals issuing from deep-ocean hydrothermal vents.
Campotohulian Eon: 1000-400 mya
The Campotohulian Eon saw the development of most of the major components of the modern Tohulian biosphere. It is often compared to the Ediacaran period on Earth, as it marked the evolution of complex multicellular life on Tohul, including the first macroscopic animal-equivalents.During the Campotohulian, the warming sun forced more water into Tohul's atmosphere, but photolysis of water in the troposphere resulted in much of it being subsequently lost to space. Ocean levels receded enough for continents to surface for the first time, and as continental and oceanic crust became more distinct, so did the range of elevation above sea level. Storms and hurricanes became more frequent, assisting the Utpalaphyta in their development of aero-planktonic lifestyles. Some Padmaphyta also evolved in this direction, but found it difficult to compete with the Utpalaphyta; the former were unable to use diffusion to excrete waste under the relatively cool atmospheric conditions, and were thus mainly relegated to the hot, dark oceans. The Phlegethonia continued to live primarily in the ocean basins, where they subsisted on decaying biomass from blooms of aquatic photosynthesizers.
Land environments during this eon were initially very wet due to constant precipitation, and were quickly colonized by Utpalaphyta. As conditions became drier, members of both Utpalaphyta and Padmaphyta were able to successfully adapt. Fossils from this period suggest that the first multicellular life on Tohul evolved around this time, appearing to have been a moss-like organism belonging to Utpalaphyta; this organism seems to have left no descendants.
As more carbon dioxide was released into the atmosphere, the surface temperature and pressure increased. The cold-adapted Utpalaphyta died out in the oceans and on the land, with the majority of surviving members being exclusively aeroplanktonic. Meanwhile, the higher ambient temperatures and millions of years of metabolic adaptations allowed the Padmaphyta to more efficiently photosynthesize under the new conditions, and as they spread and diversified they produced two new lineages, the gold-pigmented Pitamaphyta and the brown-pigmented Fuscophyta. These collectively out-competed the remaining aeroplanktonic Utpalaphyta and drove them to extinction towards the end of the Campotohulian.
Members of the Phlegethonia became increasingly able to survive throughout the oceanic water column as temperatures rose, and shifted toward becoming primarily detritivores and decomposers over hydrothermal vent feeders. Eventually, one lineage evolved multicellularity, subsequently giving rise to the animal-like kingdom Tohulanimalia along with three kingdoms of sessile saprotrophic organisms: Thermomycota ("fungoids"), Pulvispongia ("land-sponges") and Putrephyta ("putrephytes").
Some Thermomycetes developed complex tissues, giving rise to the Higher Fungoids; the rest, known as Lower Fungoids, formed aggregate communities similar to Terragen slime molds. One lineage of marine Higher Fungoids developed photosynthesis during this time, becoming the Gray Algae (Crepusculophyta); these would go extinct as the surface became darker during the Tartarean Eon.
The Tohulanimalia broadly developed in a roughly similar fashion as animals on Earth, starting out as sessile marine organisms broadly resembling corals, sponges and tunicates. These made the transition to land fairly easily due to the high humidity, and eventually developed more robust muscular systems to allow them to crawl out from their coastal tide pool habitats in search of food. Toward the end of the Campotohulian, one group of quadrilateral Tohulanimalia developed secondary bilateral symmetry, becoming the Tohulbilateria. These would become the ancestors of many of the larger and more active modern Tohulanimalia lineages, including the To'ul'hs.
Tartarean Eon: 400-300 mya
During the Campotohulian Eon, photosynthesis became possible on the surface of Tohul, but during the Tartarean the atmosphere grew so thick as to render surface-level photosynthesis almost entirely noncompetitive compared to saprotrophy feeding on the abundant organic matter falling from the higher levels of the atmosphere. This eon also saw the differentiation of Tohul's surface into its four primary modern biogeographic regions: mountains, highlands, lowlands, and ridgelands.As the surface grew hotter and darker, Padmaphyta on the surface were divided into more fungus-like heterotrophic forms closer to sea level, and holdouts of moss-like photoautotrophs on mountaintops; however, the surface soon grew too hot for either of these lineages to survive, leaving only aeroplanktonic forms in the middle and upper atmosphere. The surface-level saprotroph niche was quickly occupied by the Thermomycota.
In the realm spanning the lower atmosphere and the surface, members of the Asphodelia developed into particulate feeders, largely retaining their encrusting mold-like ancestral forms but also evolving basket-like filtering structures. One clade of Thermomycetes evolved gamete packets that used gaseous metabolic by-products for lift, allowing them to reach great heights before eventually rupturing and allowing the wind to carry their spores for hundreds of kilometers.
During this time, some lineages of microbial prokaryote-equivalents on the surface began generating methane and hydrogen in sufficient quantities to form pockets of trapped gas beneath their fibrous microbial mats, in some cases forming huge domes and bubbles that strained to be released. Some of these would break free without rupturing in the process, and be carried into the upper atmosphere. Along the way, they became inundated with various types of aeroplanktonic Elysia, which thrived off the nutrients brought from the surface. Eventually, these highly divergent groups of single-celled organisms became integrated into primitive "multi-cellular" colonial communities, with various microbes specializing into "roots" and membranous "wings" or "sails." These assemblages could control their altitude by adjusting the amount of hydrogen in their gasbags, rising to get more sunlight for photosynthesis or sinking to gather more nutrients. Although relatively simple compared to modern cloudforests, these symbiotic communities represented the early stages of a process that would eventually lead to the evolution of cloudforests.
On the surface, the Phlegethonia diversified further. The constant rain of "biological snow" from the middle and upper atmospheric levels enriched the soil and water, leading to massive growths of aquatic Fungoids. Some of these evolved into forms resembling a combination of Earth sea-fans and kelp, with holdfasts that kept them firmly planted in the seabed. In response to the decreasing light levels, the various groups of surface-living Tohulanimalia gradually became more reliant on non-visual senses, such as smell and echolocation; by the end of the Tartarean, the majority had become nearly or completely blind.
Caelosilvanic Eon: 300 mya-present
The Tartarean Eon transitioned into the Caelosilvanic around 300 million years BT, when various lineages of sky-algae and other organisms began to aggregate into floating ecosystems, similar in many ways to coral reefs, which today are known as "cloudforests." These have since become a key feature of the modern Tohul biosphere, and give the Caelosilvanic Eon its name.The Balloon Fungi continued to diversify during this eon. One lineage, the Sky-Lanterns (Volucernidae), evolved bioluminescence to attract flying animal-equivalents, which would then become coated in spores that they would take back to their cloudforest roosts, allowing the fungi to colonize the rich soil trapped in the floating masses. In one lineage, the sporozoan stage of the life cycle became dominant, developing into motile animal-like forms capable of powered flight. The most basal of these lineages, the Saganoptera ("Sagan's Fliers", so named for their striking resemblance to hypothetical Jovian organisms proposed by the Information Age scientist and fabulist Carl Sagan) developed a primitive form of filter-feeding, using their four membranous wings as traps for aeroplankton.
Members of the Asphodelia also became more prominent in the middle atmospheric zones, dangling sticky tentacles below their gas-filled lifting bodies to trap aeroplankton and larger flying organisms that they would then digest externally. These Aeromedusoids underwent a significant adaptive radiation in the early Caelosilvanic, developing into myriad forms.
On the ground, the Tohulbilateria continued to diversify. The hard-bodied Tohularthropods appeared during the early Caelosilvanic, evolving to occupy a vast array of niches and taking on a multitude of forms. Several lineages developed powered flight, and as the cloudforest ecosystems became more prevalent and complex, some of these colonized the middle atmosphere by hitching rides on rising cloudforest biomass, eventually giving rise to the Skybeetles (Caelocoleopteroidea), a group adapted to the colder temperatures of the cloudforest zone. Another group, the Rootworms (Filovermidae) lost their hard shells and bilateral symmetry entirely and adopted bizarre, branching bodyplans somewhat resembling the root systems of Terragen plants, using long filaments to maximize their surface area and absorb nutrients from the water and soil.
Yet another subset of Tohulbilateria evolved centralized nervous systems, giving rise to the Tohulichordates. One group of soft-bodied amphibious Tohulichordates broadly resembling large tardigrades or onychophorans split into fully aquatic and terrestrial lineages, giving rise to the Blindfish (Caecipisciformes) in the seas and the Slug-bugs on the land. The Blindfish subsequently went on to become one of the largest and most successful groups of aquatic Tohulanimalia, occupying a wide variety of niches. Meanwhile, the Slug-bugs diversified into the hard-shelled burrowing Shovelheads (Cephalofossoria) and the eight-legged Tohularachnida, which further split into the subterranean Mole Bugs (Talpipoda) and the surface-living Gliding Spiders (Alataranea); some of the latter would subsequently hitch rides on rising cloudforests, eventually adapting to the colder temperatures and becoming the Flying Monkeys (Xenolemuridae).
Around 225 million years BT, one lineage of Slug-bugs developed the ingenious strategy of cultivating and tending to hydrogenic microbial mats, shaping their gas pocket-enclosing biofilms into lifting bodies which they would then cut free of the rest of the mat and ride into the middle and upper atmosphere. Some of these went a step further, forming a symbiotic partnership with hydrogen-producing microbes such that they would grow gas-bubble biofilms directly on their dorsal surfaces, effectively integrating the lifting bodies into their anatomy. Some of these Float-bugs further integrated these microbial symbionts into dedicated buoyancy organs supported by a rigid exoskeleton, becoming the Zeppelin Bugs (Hindenburgia). Others lost their exoskeletons, becoming soft-bodied, lightweight and relatively fragile creatures that mostly drifted on the wind. Some of these developed wing-like appendages enabling powered flight, becoming the Skyfish (Aeropisciformes). Over the following 125 million years these would diversify into numerous different clades, including the small jet-propelled Nitrobats (Flatusfugidae) and the enormous filter-feeding Skywhales (Caelocetoidea).
The Tohulcephalochordata appeared around 210 million years BT, with the earliest members being aquatic four-armed cephalopod-like creatures. One lineage of these took on a variety of fast-moving pelagic forms broadly analogous to Terragen fish and squid, while a later group gave rise to the large, predatory bottom-dwelling Krakenoids, which subsequently went extinct around 20 million years BT; one subgroup of these, the Cthulhunids, were the largest animal-equivalents by mass ever to have evolved on Tohul. Another offshoot of the Tohulcephalochordates evolved articulated internal skeletons, becoming the four-legged terrestrial Tohultetrapoda. The Tohultetrapods diversified into many new lineages, with some of the most prominent genera including the Shur'roos'hur (Lututherium spp.), H'shass'thee ("Spockbats", Deinoparvichiropteryx spp.), and the To'ul'hs (To'ul'h sapiens).
Around 50 million years BT, one lineage of Skybeetles, the H'thik ("Locust-Beetles", Khepridae spp.), became re-adapted to the hot surface conditions and began to colonize the ground, adopting a life-cycle in which their larvae remained in the upper atmosphere until maturation and descended to the surface as adults. Traveling in massive swarms and indiscriminately consuming both motile and sessile organisms alike, the Locust-Beetles remain a major component of the modern Tohul surface ecosystem.
During the roughly 100 millennia of To'ul'h civilization prior to Terragen contact, the majority of To'ul'h societies employed relatively conservative and low-impact agricultural practices, a choice born partly out of necessity due to the low primary productivity of Tohul's surface environments. However, during this time a number of short-lived but highly-active civilizations intermittently arose which caused long-lasting disruptions to Tohul's surface and atmospheric ecosystems; among the most notable of these were the various "skywhaler" cultures that arose between 10,000 and 7,500 BT, which sought to exploit the rich biota of the middle and upper atmosphere using dirigibles and floating resource-harvesting platforms. Though the long-term environmental impact of these cultures was relatively small compared to the effects of Terragen civilization on Earth's biosphere following the Industrial Revolution, their activities nonetheless resulted in the extinction or drastic depopulation of numerous species, including many species of surface and aerial megafauna. Most of the affected taxa have since been lazurogened or otherwise rehabilitated through the efforts of O'hoth'so'toh following Tohul's appropriation by the Caretaker God in 4645 AT, about 814 years after initial Terragen contact.
Related Articles
- Clade (evolution)
- Cladogram (evolution) - Text by M. Alan Kazlev
Branching diagram that depicts divergence of the analysed taxa from their common ancestors. It shows the distribution and origins of unique shared characteristics (synapomorphies and autapomorphies), as well as their monophyly. A testable hypotheses of phylogenetic relationships. - Eden Institute of Xenoscience
- Evolution (biology)
- Evolutionary Tree - Text by M. Alan Kazlev
Phylogenetic or cladistic diagram tracing ancestry-descent, branching, cross-links of genetic/informational and morphotypic exchange, and other factors in order to provide a complete and usually multi-parameter diagram of the evolutionary history of any taxon. A beautiful collection of evolutionary trees can be seen in the great Phylogeny Orbitals of Darwinia (NuiHibbert Sector, Zoeific Biopolity). - Phylogeny - Text by M. Alan Kazlev
The science and art of tracing family trees of ancestry and descent, whether regarding the evolution of species over millions of years, or of terragen clades or genetic houses over thousands of years. - Phylum - Text by M. Alan Kazlev
In the Linnean taxonomic classification, this is the second highest grading, and designates a phylum is a major group of organisms sharing a similar bodyplans. A phylum contains one or more classes. A group of similar phyla forms a Kingdom. - Shur'rooss'hur
- Skywhale (To'ul'h species)
- Spockbat
- Taxon - Text by M. Alan Kazlev
Formal supra-individual rank of biological or artificial organisms - e.g. species, phylum, etc. (pl. taxa). - Tohul (To'ul'h Homeworld)
- Tohul Fauna
- Tohulian Worlds
- Xenobiochemistry
- Xenobiologist
- Xenobiont
- Xenobiota - Text by M. Alan Kazlev
The totality of non-terragen life on a specific planet or non-terragen ecosystem. - Xenologist
- Xenophotosynthesis
Appears in Topics
Development Notes
Text by Andrew P.
ProxCenBound, Immanuelle, The Astronomer, Dangerous Safety , James Rogers, Todd Drashner, Steve Bowers
Initially published on 09 July 2014.
expansion published in 18 April, 2025 replacing the article "To'ul'h Prime Phylogenetic Tree" (09 July 2014) by Andrew P, with additional material by Steve Bowers; from ideas by Stephen Inniss and Anders Sandberg
ProxCenBound, Immanuelle, The Astronomer, Dangerous Safety , James Rogers, Todd Drashner, Steve Bowers
Initially published on 09 July 2014.
expansion published in 18 April, 2025 replacing the article "To'ul'h Prime Phylogenetic Tree" (09 July 2014) by Andrew P, with additional material by Steve Bowers; from ideas by Stephen Inniss and Anders Sandberg